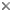The three subfamilies of Fabaceae (Caesalpinioideae, Mimosoideae, Papilionoideae) all contain species that form root nodules as described by Sprent (2001) in Nodulation in Legumes. In Mimosoideae, all genera appear to nodulate, but not all species do; for example, within tribe Acacieae, non-nodulating species are seen only in the subgenus Aculeiferum (Sprent, 2001; pp. 49-58). Papilionoideae has the highest proportion of nodulating species, and produce all known types of nodules. Some papilionoid tribes, such Dipterygeae with three genera, lack the ability to nodulate, while larger tribes such as Dalbergieae, Swartzieae, and Sophoreae contain both non-nodulating and nodulating genera (Sprent, 2001; pp. 63-106). In Caesalpinioideae, there are reports of nodulation by members of all tribes but the strongest evidence of nodulation appears confined to members of tribes Caesalpinieae and Cassieae (Sprent, 2001; pp. 40-48).
"Rhizobia" is the name given to the group of genera of alpha-proteobacteria (family Rhizobiaceae) which includes all of the nitrogen-fixing species that produce nodules with legumes, Allorhizobium, Azorhizobium, Bradyrhizobium, Mesorhizobium, Phyllobacterium, Rhizobium, and Sinorhizobium, as well as the plant pathogen Agrobacterium (Sprent, 2001; pp. 15-18). Rhizobia and their legume host must recognize each other for nodulation to begin. Different species of rhizobia are specific to their host plants, but often can infect more than one species of plant. Additionally, a single species of plant can be infected by more than one species of bacteria. Sprent (2001; pp. 52) gives the example of Acacia senegal, which is nodulated by seven species of rhizobia from three different genera. One of the major defining characteristics between the genera of rhizobia is the rate at which they grow (whether they are fast or slow growing). Those that are fast growing synthesize acidic products and those that are slow growing sythesize alkaline products (Sprent, 2001; p. 14). These differing nitrogen products are also specific to nodule shape. Root nodules are classified as indeterminate, which are cylindrical and often times branched, producing amide products, and determinate, which are spherical with prominent lenticels, producing ureide products. Indeterminate nodules are characteristic of peas and alfalfa, plants of temperate regions, and determinate nodules are characteristic of soybeans and similar plants, plants of tropical and sub-tropical regions (Sprent, 2001; p. 35). Indeterminate nodules are the only type found in Caesalpiniodeae and Mimosoideae and the most common type in Papilionoideae (Sprent, 2001; p. 32).
As a result of the variety of ways the plant is infected by rhizobia, there are different types of nodules. Roots can be infected via the root hairs, damaged epidermal tissues, or intact epidermis. During infection via the root hairs, the plant and rhizobia must first recognize each other for rhizobia colonization to occur. Colonization occurs on the root surface and the rhizobia attach to the root hairs. The bacteria synthesize compounds called 'nod' factors that initiate the nodulation process in the plant (Sprent, 2001; p. 26). Upon recognition of the proper rhizobia, the growing root hairs curl and form a pocket for the rhizobia. This allows the bacteria to enter the plant through the root hair cell wall. The plant builds pathways, called infection threads, around the rhizobia allowing them to move between cells. With the production of the infection threads, nodules begin to form in the root hairs (Sprent, 2001; pp. 28-30). Rhizobia infect the root hair to form indeterminate nodules just before tip growth, while those of determinate nodules infect at the beginning of growth (Sprent, 2001; p. 35). Some species of plants house the bacteria in fixation threads similar to infection threads rather than in separate nodules. This variety of nodulation is thought to be a result of plant and bacteria species that are not completely compatible for root nodulation or a primitive form of nodulation (Sprent, 2001; p. 32). Fixation threads are observed in Caesalpinioideae and Papilionoideae, but not in Mimosoideae (Sprent, 2001; p. 34).

bm_27-03.250a.jpg)

thomas-oates_f1.250a.jpg)
Root nodules from bur clover, Medicago spp., "stem" nodules on Sesbania rostrata, and soybean (Glycine max) root nodules.
Rhizobia can infect their hosts through damaged epidermis in addition to infection through the root hairs. In the formation of root nodules via the root hair, the rhizobia must induce the root hairs to curl and form a pocket that allows them to enter the root cells. In wound or crack infections, the rhizobia can enter the plant directly without these preliminary processes. In this instance, nodules often form close to the stem of the plant, so are called stem nodules. Stem nodules are essentially nodules that are initiated at lateral root primordia, most often because the plant’s roots are water logged; this form of nodulation is often observed in Neptunia and Sesbania (Goormachtig et al., 2004). Lupin nodules are distinct, characterized by collar-like nodules initiated by wound infections and most prominent around the taproot (Sprent, 2001; p. 37). Rhizobia can initiate infection through undamaged cells as described by Faria et al. (1988) in Mimosa scabrella; they observed that Mimosa scabrella is infected by rhizobia that digest the root in order to enter the plant’s cells (Sprent, 2001; p. 38).