Nodosauridae
Kenneth Carpenter


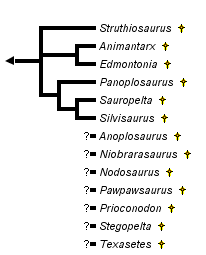

This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Our modern concept of ankylosaurs has been slow in development due to the mistaken belief that all ankylosaurs were characterized by spines projecting from the body and a large club on the end of the tail. Based on the work of Walter Coombs (1978a) and Carpenter (1982, 1984), nodosaurid ankylosaurs are now known to be characterized by outwardly projecting spines on the neck and shoulders (Fig. 1), while ankylosaurids have a tail club.
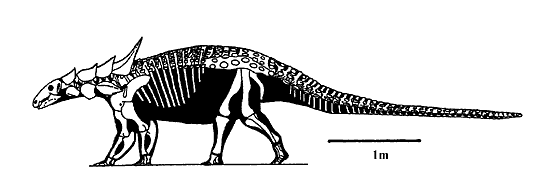
Figure 1: Skeleton of Sauropelta, the best known nodosaurid from the Early Cretaceous of North America. Characteristic features of the nodosaurids include the absence of a tail club, long spines on the neck and shoulder, bent ischium (pelvic bone seen protruding between legs and tail).
The earliest definitive nodosaurid is Sauropelta from the lower part of the Cloverly Formation, which is late Aptian in age (it is also present in the overlying Albian portions of the Cloverly Formation). Older specimens have been referred to the Nodosauridae (e.g., Sarcolestes leedsi from the Callovian of England), but these are either nomen dubia or have been referred to the family Polacanthidae (Carpenter, 2001). Sauropelta is one of the largest nodosaurids, with a length of 5.5 m (Fig. 1). Interestingly, such large nodosaurids are unknown from Cretaceous deposits of the European Archipelago, leading Pereda-Suberbiola (1992) to refer to the European nodosaurids as "dwarf insular forms." Nodosaurids appear in Antarctica by the Campanian (83-71 mya) (Gasparini et al., 1996), probably through South America because Campanian nodosaurids are also known from there (Coria and Salgado, 2001). Nodosaurids are not known from the continental deposits of China and Mongolia, although a possible specimen has been reported from the Cenomanian marine deposits of northern Japan (Hayakawa and Carpenter, 1996).
Galton (1980) has inferred a European origin for the Nodosauridae because of Middle Jurassic specimens referred to that family, and because, under the old bipartite classification of ankylosaurs, the Nodosauridae was the only family present in Europe. With the recognition of a second family in Europe, the Polacanthidae, and the dubious nature of the Middle Jurassic specimens, there is less certainty that nodosaurids originated in Europe.
The extinction of nodosaurids in North America and Europe appears to have occurred roughly at the same time in the late Maastrichtian (~66-67 mya), but well before the end of the Maastrichtian (65 mya) and the end of the dinosaurs. Carpenter and Breithaupt (1986) documented an absence of specimens above about the middle of the Lance and Hell Creek formations. Such an absence is believed to be real because of the thousands of person hours spent looking for fossils stratigraphically higher on both sides of the K-T boundary by various individuals. The extinction in both North America and Europe may have been due to a loss of coastal habitats coincidental with the regression of many shallow continental seas that occurred globally at that time (see coastal maps in Smith et al., 1994).
Characteristics
Nodosaurid skeletons are conservative making it difficult to identify some individual bones to a particular genus. The skull is typically pear-shaped in top view, with impressions of large scales, especially on the skull roof (Fig. 2). Typically, there is a single scale impression between nostril openings, and a narrow scale impression along the posterior edge of skull. The palate has an hourglass-shape, with the maxillary tooth row narrowest in the middle. The cutting edge of the premaxilla is connected to the maxillary tooth row by a low ridge, thus separating the palatal area from the cheek emargination. At the base of the skull, the basipterygoid processes usually consist of a pair of rounded, rugose stubs. At the rear of the skull, the hemispherical occipital condyle is composed of the basioccipital only. Furthermore it is set off from the braincase by a short neck that is angled downwards at about 50º. The jugal and postorbital "horns" are small, if present, and usually rounded. On the lower jaw, the coronoid process is very tall.

Figure 2: Skull of Edmontonia in side (left) and top (right) views. The characteristic features of the nodosaurid skull that may be seen include: length greater than width, large symmetrically arranged scale impressions into the skull bones (ornamentation).
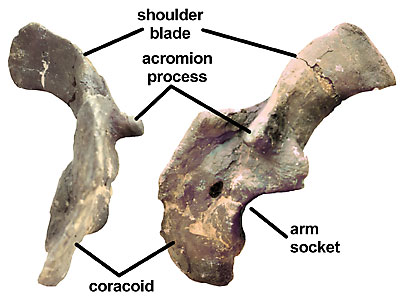
Figure 3: Shoulder girdle of Panoplosaurus showing the characteristic ridge-like acromion of nodosaurids.
The shoulder blade, or scapula (Fig. 3), has a distinct ridge-like process, the acromion, above the glenoid, or arm-socket, for the M. scapulohumeralis anterior (Coombs, 1978b). The shape of the acromion and its location relative to the glenoid is taxonomically distinct at the generic level. The coracoid attached to the scapula is large and elongate. In the pelvis, the ischium is ventrally bent near mid-length (Fig. 1). Finally, the armor of nodosaurids is generally thick and solid, rather than the thin-wall or hollow base seen in polacanthids and ankylosaurids.
Discussion of Phylogenetic Relationships
Phyletic analysis of the nodosaurids is very difficult owing to the incompleteness of most of the specimens. Several taxa are known only from single, incomplete holotype specimens. Only two taxa, Sauropelta and Edmontonia are known from enough overlapping specimens to be reconstructed with confidence. Only two previous phyletic analyses of nodosaurid genera have been presented. The earliest was presented by Coombs and Maryanska (1990):
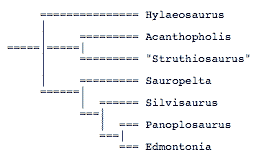
This analysis includes taxa that have been moved to the family Polacanthidae (e.g., Hylaeosaurus) or are nomen dubium (e.g., Acanthopholis). They also used characters of dubious value (e.g., increase in body size), or used the characters inconsistently (e.g., presence or absence of premaxillary teeth). Furthermore, Hylaeosaurus is placed as the most plesiomorphic taxon without justification.
The second analysis, by Lee (1996), used only cranial characters, and thus is of limited value:
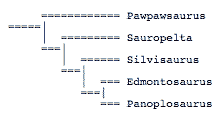
It contains fewer taxa than another presented in Carpenter (2001), which uses both cranial and postcranial characters.

The more inclusive analysis establishes two major, unnamed nodes. However, the analysis also includes taxa for which more than 50% of the characters are unknown. A more restrictive analysis using only taxa with 50% or more of the characters known does not support these two nodes and even the position of the taxa is different:
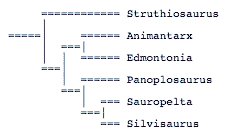
Paleobiology
Nodosaurid ankylosaurs appear to have been restricted to low coastal environments of North America and Europe, as well as the upland environments of North America. Paleoenvironmental reconstructions of these areas, based on sedimentology and fossil plants, indicate warm, wet subtropical coastal plains, and strongly seasonal (wet-dry) uplands. Nodosaurid remains are more common in the coastal plain environments of the Late Cretaceous, than in the upland environments.
The muzzle of nodosaurids is narrow as compared with ankylosaurids. Based on analogies with modern ungulates, the narrow muzzle suggests selective feeding. The teeth are adapted for vegetation and show some tooth-to-tooth wear. The presence of cheek plates, formed within the skin, indicates nodosaurids had a fleshy, probably non-musculature, cheek. This cheek and the tooth wear suggest that nodosaurids chewed their food (oral processors).

Figure 4: Front view of the mounted skeleton of Edmontonia (American Museum of Natural History), Specimen is from the Dinosaur Park Formation, Dinosaur Provincial Park, Alberta, Canada. The outward projecting spines make the individual look wider and larger than it really was. This indicates that display, rather than defense, may have been a major feature of armor.
Nodosaurids have three rows of keeled plates on the neck, whereas both polacanthids and ankylosaurids have but two. The armor of nodosaurids shows the greatest variation around the head (Fig. 1 and 4). Most of the armor consists of long spines or spikes, many of which project backwards where they could not adequately serve as weapons. Instead, the armor may have played an important role in visual display. Edmontonia does have some spines that project laterally (Fig. 4), which may have made the animal appear wider, hence larger than it really was. It also has a bifurcated forward projecting spike on each shoulder that could serve as a weapon. However, since the spike is apparently larger in males than females (Carpenter 1990), it might have also been used in shoving matches among males (title figure).
References
Carpenter, K. 1982. Skeletal and dermal armor reconstruction of Euoplocephalus tutus (Ornithischia: Ankylosauridae) from the Late Cretaceous Oldman Formation of Alberta. Canadian Journal of Earth Sciences 19:689-697.
Carpenter, K. 1984. Skeletal reconstruction and life restoration of Sauropelta (Ankylosauria: Nodosauridae) from the Cretaceous of North America. Canadian Journal of Earth Sciences 21:1491-1498.
Carpenter, K. 1990. Ankylosaur systematics: example using Panoplosaurus and Edmontonia (Ankylosauria, Nodosauridae). Pp. 282-298 in Carpenter, K., and Currie, P. (eds.), Dinosaur Systematics: Perspectives and Approaches. Cambridge University Press, New York.
Carpenter, K. 2001. Phylogenetic Analysis of the Ankylosauria. Pp. 455-483 in K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
Carpenter, K., and Breithaupt, B. 1986. Latest Cretaceous occurrence of nodosaurid ankylosaurs (Dinosauria, Ornithischia) in western North America and the gradual extinction of the dinosaurs. Journal of Vertebrate Paleontology 6:251-257.
Coombs, W. 1978a. The families of the ornithischian dinosaur order Ankylosauria. Journal of Paleontology 21:143-170.
Coombs, W. 1978b. Forelimb muscles of the Ankylosauria (Reptilia: Ornithischia). Journal of Paleontology 52:642-658.
Coombs, W., and Maryanska, T. 1990. Ankylosauria. Pp. 456-483 in D. Weishampel, P. Dodson and H. Osmolska (eds.), The Dinosauria. University of California Press,Berkeley.
Coria, R.A. and Salgado, L. 2001. South American Ankylosaurs. Pp. 159-168 in K. Carpenter (ed.). The Armored Dinosaurs. Indiana University Press, Bloomington.
Galton, P.M. 1980. Armored dinosaurs (Ornithischia: Aankylosauria) from the Middle and Upper Jurassic of Englnd. Geobios 13: 825-837.
Gasparini, Z., Pereda-Suberbiola, X., and Molnar, R. 1996. New data on the ankylosaurian dinosaurs from the Late Cretaceous of the Antarctic Peninsula. Memoirs of the Queensland Museum 39:583-594.
Hayakawa, H. and Carpenter, K., 1996. First occurrence of nodosaurid ankylosaurs in Aasia. Palaeontological Society of Japan Abstracts: 98.
Lee, Y.-N., 1996. A new ankylosaur (Dinosauria: Ornithischia) from the Paw Paw Formation (Late Albian) of Texas. Journal of Vertebrate Paleontology 16:232-245.
Pereda-Suberbiola, J., 1992. A revised census of European Late Cretaceous nodosaurids (Ornithischia: Ankylosauria): last occurrence and possible extinction scenarios. Terra Nova 4:641-648.
Smith, A.G., Smith, D.G., and Funnell, B.M. 1994. Atlas of Mesozoic and Cenozoic coastlines. Cambridge University Press, Cambridge.
Title Illustrations

| Scientific Name | Edmontonia |
|---|---|
| Location | North America |
| Comments | Life restoration. Two males are shown in a shoving contest of strength. |
| Specimen Condition | Fossil -- Period: Late Cretaceous |
| Sex | Male |
| Copyright |
© 1983 Brian Franczak

|
About This Page
Thanks to Katja Schulz, The Tree of Life Project, for her asistance in getting these pages designed and online, and to Brian Franczak for permission to use the image of Edmontonia.
All other illustrations are by the author, copyright © 2000 Kenneth Carpenter.
Denver Museum of Natural History, Denver, Colorado, USA
Page copyright © 2000
 Page: Tree of Life
Nodosauridae.
Authored by
Kenneth Carpenter.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Nodosauridae.
Authored by
Kenneth Carpenter.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 22 December 2000
- Content changed 10 January 2002
Citing this page:
Carpenter, Kenneth. 2002. Nodosauridae. Version 10 January 2002 (under construction). http://tolweb.org/Nodosauridae/15772/2002.01.10 in The Tree of Life Web Project, http://tolweb.org/