Dermaptera
Earwigs
Fabian Haas



- Archidermaptera
 (non-monophyletic)
(non-monophyletic) - Hemimerina
- Arixenina
- Forficulina
Introduction
Earwigs or Dermaptera constitute a comparatively small order of insects comprising about 1800 species. Their scientific name refers to the leathery fore wing (dermatos: skin, pteron: wing, both words of Greek origin) and was given by de Geer in 1773. However, he used the name for all orthopteroids. It was Kirby in 1815 who introduced the name in the modern sense.
Dermaptera are elongate and slender insects with a prognathous head. If not wingless, the thorax bears two pairs of wings of which the first one is small and leathery, called tegmina. The second pair is large and membraneous (see fig. below), almost semi-circular, and, at rest, folded underneath the tegmina in a complicated manner. The abdomen of extant Dermaptera is highly movable with a pair of unsegmented (not always forceps-like) cerci at its posterior end. The body length ranges from approximately 4 to 80 mm including cerci. Most Dermaptera are uniformly coloured brown or black, sometimes with a light brown or yellow pattern; other colours, like metallic green are real exceptions. They are distributed throughout the world (except the polar regions), with greatest diversity in the tropics.
Not very much is known about the natural history of the Dermaptera as a whole, with the species studied in some detail being mostly European. So "the earwig" is Forficula auricularia, the common earwig; other well studied species are Labidura riparia, the striped earwig and Anisolabis maritima, the seaside earwig.
Most species are omnivorous but predominantly phytophagous or predacious species are also known. Some species live on decaying material. The Hemimerina feed on scurf and fungi on the skin of giant rats without harming them. The Arixenina feed on skin gland secretions of bats and occasionally on dead insects.
Dermaptera are thigmotactic (searching close contact to surfaces). They frequent (humid) crevices of all kinds: you can find them under bark, between leaves and under stones. Probably, earwigs also creep in human ears just taking them for humid and warm crevices to hide. This behaviour probably gave rise to the belief that they penetrate the tympanum and lay eggs in the brain, which is utter nonsense. Therefore their name in colloquial speech often refers to the ear, like in Danish, Dutch, English, French, German, Russian and Swedish, or to the forceps, like Italian, Finnish, Portuguese and Spanish. The behaviour of Dermaptera is complex and the cerci seem to play an important role. They are used to open the wings, to capture prey and for defence. Females show maternal care.
The development is hemimetabolous. The larvae (4 to 5 instars) resemble the adult apart from having only wing buds. However, in wingless species the larvae are often difficult to distinguish from the adults. The larva's cerci are simple, almost straight and resemble those of the female. Vivipary occurs in all Hemimerina and Arixenina but is not known from the Forficulina, which are generally oviparous.
The fossil record of the Dermaptera begins in the Jurassic (Carpenter, 1992), about 208 million years ago and comprises about 70 specimens. The earliest fossils are very similar to Recent species but adults have segmented cerci. No fossil Hemimerina and Arixenina are known. The probable stem group of the Dermaptera are the Protelytroptera (Carpenter, 1992; Hennig, 1969). These insects, which resemble modern Blattodea or Cockroaches, are known from the Permian (about 290 MYA) of North America, Europe and Australia (Carpenter, 1992). There are no fossils from the Triassic when the morphological changes from Protelytroptera to Dermaptera took place.
The Dermaptera are often considered a neglected group. However, there are a number of monographs and regional faunas with a large general section, which give a good introduction to their biology: Albouy & Caussanel (1990), Beier (1953, 1959), Bey-Bienko (1936), Bormans & Krauss, (1900), Brindle (1972), Burr (1910), Carpenter (1992), Chopard (1938, 1949), Günther & Herter (1974), Harz (1960), Rentz & Kevan (1991), Sakai (1970-1996), Steinmann (1986, 1989, 1990, 1993).
Characteristics
Even though Dermaptera are easily recognized, it is not easy to find synapomorphic characters for the whole order. Most striking are unsegmented, forceps-like cerci, however, they are not forceps-like in the Hemimerina and Arixenina. Furthermore, unsegmented cerci occur in many orthopteroids.
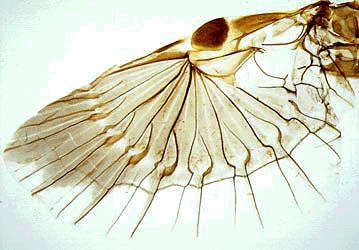
Unfolded wing of Allodahlia scabriscula, Forficulina: Forficulidae
The morphology of their tegmina and hind-wings could constitute a synapomorphy but the Hemimerina and Arixenina are wingless. Maternal care is remarkable but only a few species have been studied. However, the occurence of maternal care in the very primitive genus Diplatys and in the highly derived genus Forficula suggests that this is a common trait for all Forficulina. There are almost no observations concerning maternal care in Hemimerina and Arixenina. In female Forficulina and Hemimerina, the tergites of abdominal segment 8 and 9 are narrow, fused and covered by the tergite of segment 7. However, this is not true for the Arixenina (Giles, 1963). The tarsi have three segments in all Recent suborders. In all Dermaptera, the first abdominal sternite is lacking and the tergites and sternites are overlapping laterally. A cytological trait, holocentric chromosomes, probably is the most convincing synapomorphy. It is found in all Recent suborders and is unique in orthopteroid insects (Rentz & Kevan, 1991).
The Suborders of Dermaptera
The Dermaptera are divided into four suborders.
The Archidermaptera comprise ten fossil species from the Jurassic with segmented cerci in adults and four or five segmented tarsi (Bey-Bienko, 1936). Both character states are considered primitive for Dermaptera and used as diagnostic features.
The Forficulina are by far the largest group comprising about 1800 species in 180 genera, including the common earwig and the striped earwig. The body shape of these species is characteristic for the whole suborder. The species are omnivorous, phytophagous or predacious. The adult cerci are unsegmented and forceps-like. The larval cerci are unsegmented except in two primitive groups. If not wingless, the hind wings are complexly folded in a unique way, and covered by tegmina.
The viviparous Hemimerina comprise ten species in one genus living epizoically on giant rats in tropical Africa. They are wingless, blind, about 10 mm long excluding the filiform cerci, and possess short and stout legs. Their body is streamlined and smooth for rapid movement through the fur.
The viviparous Arixenina comprise five species in two genera and live on bats in the Malayan-Philippinian region. They are wingless, almost blind, possess straight rod-like cerci and have long and slender legs.
Discussion of Phylogenetic Relationships
The phylogenetic relationships of the four suborders are not well understood. The suborders are usually described separately because they are cleary different in morphology and natural history. The discussion of their relationships have been further complicated because no phylogenetic system has been available for the Forficulina. Therefore, the evolution of characters remained obscure, which otherwise could help to interpret the characters of the highly derived Hemimerina and Arixenina. The discussion of their relationships was largely based on considerations of similarities and differences between the groups and not on a cladistic analysis of characters.
The systematic position of Hemimerus was subject to considerable change in the first years after its discovery. It is beyond the scope of this page to give a detailed account of the discussion but the topic was whether or not Hemimerus should be placed in a separate order or included in the Dermaptera (for a detailed account see Giles (1963)). The subordinal rank was given by Burr (1911). The genus Arixenia was first described by Jordan (1909). Again, Burr (1911) erected a new suborder, the Arixenina. So, the widely accepted view (for example Sakai (1982)) that the order Dermaptera consists of three Recent suborders is based on Burr (1911).
In 1985, Popham re-assessed the relationships of the Dermapteran suborders and families using cladistic methods. His phylogenetic system is mainly based on characters of the genitalia:
=== Hemimerina
|
| === Pygidicranidae
==| |
| | === Carcinophoridae (Anisolabididae)
===| |
| ===| === Arixeniidae
| | ===|
===| === Labiidae (Spongiphoridae)
|
| === Labiduridae
| |
======| === Chelisochidae
===|
=== Forficulidae
The Hemimerina are still regarded as a suborder, whereas the Arixenina are treated as the probable sistergroup of Spongiphoridae, accordingly called Arixeniidae. Popham's view has not been generally accepted. He split traditional groups, like the Eudermaptera (Spongiphoridae+Chelisochidae+Forficulidae) and regarded the Carcinophoridae to be wingless. However, there is a fair number of fully winged species in this family, among them Carcinophora americana. Other characters have been criticized by Haas (1995), Willmann (1990) and Günther & Herter (1974) referring to Popham (1965). Steinmann (1986) regards Dermaptera to consist of only Forficulina. However, this view is not widely shared.
The position of the extinct Archidermaptera has hardly been discussed. Unsegmented adult cerci and four or five segmented tarsi are generally regarded as plesiomorphic character states. Willmann (1990) presented the first phylogenetic study. He concluded that the Archidermaptera should be regarded as sistergroup of the remaining suborders.
References
Albouy, V. & Caussanel, C. (1990) Dermapt?res ou Perce-oreilles. Faune de France 75. F?d?ration Fran?aise des Soci?t?s de Science Naturelle, Paris. 245pp.
Beier, M. (1953) Dermaptera - Ohrw?rmer. Biologie der Tiere Deutschlands, 26, 169-231pp.
Beier, M. (1959) Ordnung: Dermaptera. Ed. by H. Weber: Bronn's Klassen und Ordnungen des Tierreichs Bd V, Abt. III Lief. 3. Akademische Verlagsgesellschaft, Leipzig. 455-586.
Bey-Bienko, G.J. (1936) Dermapt?res. Ed. by S. A. Sernov and A. A.Stackelberg: Faune de l'URSS L'Academie des Sciences de l'URSS, Moskau, Leningrad. 239pp.
Bormans, A. & Krauss, H. (1900) Forficulidae und Hemimeridae. Ed. by Das Tierreich 11. Friedl?nder und Sohn, Berlin. 142pp.
Brindle, A. (1972) Insects of Micronesia. Insects Micronesia, 5, 97-171.
Burr, M. (1910) The Fauna of British India: Dermaptera. Taylor and Francis, London. 217pp.
Burr, M. (1911) Genera Insectorum. Ed. by P. Wytsman: 122, 1-112.
Carpenter, F.M. (1992) Superclass Hexapoda. Ed. by R. L. Kaesler: Treatise on Invertebrate Paleontology Part R Arthropoda 4. The Geological Society of America, Boulder, Colorado. 143-153pp.
Chopard, L. (1938) La biologie des Orthopt?res. Ed. by P. Lechevalier: Encyclop?die Entomologique S?rie A 20. Paris. 541pp.
Chopard, L. (1949) Ordre de Dermapt?res. Ed. by P. P. Grass?: Traite de Zoologie IX. 745-770.
Colgan, D.J., G. Cassis and E. Beacham. 2003. Setting the molecular phylogenetic framework for the Dermaptera. Insect Systematics and Evolution 34(1): 65-79.
Engel, M.S. and D.A. Grimaldi. 2004. A primitive earwig in Cretaceous amber from Myanmar (Dermaptera: Pygidicranidae). Journal of Paleontology 78(5): 1018-1023.
Giles, E.T. (1963) The comparative external morphology and affinities of the Dermaptera. Transactions of the Royal entomological Society of London, 115, 95-164.
Guillet, S. and M. Vancassel. 2001. Dermapteran life-history evolution and phylogeny with special reference to the Forficulidae family. Evolutionary Ecology Research 3(4): 441-447.
G?nther, K. & Herter, K. (1974) Dermaptera. Ed. by J.-G. Helmcke, D. Starck and H. Wermuth: Handbuch der Zoologie IV. 2. H?lfte, 2. Teil 11. Gruyter, Berlin. 158pp.
Haas, F. (1995) The phylogeny of the Forficulina, a suborder of the Dermaptera. Systematic Entomology, 20, 85-98pp.
Haas, F. and S. Gorb. 2004. Evolution of locomotory attachment pads in the Dermaptera (Insecta). Arthropod Structure and Development 33(1): 45-66.
Haas, F. and K. D. Klass. 2003. The basal phylogenetic relationships in the Dermaptera. In K. D. Klass (ed.): Proceedings of the First Dresden meeting on insect phylogeny: "Phylogenetic Relationships within the Insect Orders" (Dresden, September 19-21, 2003). Entomologische Abhandlungen 61:138-142.
Haas, F. and J. Kukalova-Peck. 2001. Dermaptera hindwing structure and folding: New evidence for familial, ordinal and superordinal relationships within Neoptera (Insecta). European Journal of Entomology 98(4).
Harz, K. (1960) Geradfl?gler oder Orthopteren. Ed. by M. Dahl and H.Bischoff: Die Tierwelt Deutschlands 46. G. Fischer, Jena. 214-229pp.
Hennig, W. (1969) Die Stammesgeschichte der Insekten. Kramer, Frankfurt a.M. 436pp.
Jarvis, K.J. and M.F. Whiting. 2004. The phylogeny of Dermaptera: Molecular evidence. Cladistics 20(1): 86.
Jordan, K. (1909) Description of a new kind of apterous earwig, apparently parasitic on a bat. Novit. zool., 16, 313-326.
Kamimura, Y. 2004. In search of the origin of twin penises: Molecular phylogeny of earwigs (Dermaptera: Forficulina) based on mitochondrial and nuclear ribosomal RNA genes. Annals of the Entomological Society of America 97(5): 903-912.
Popham, E.J. (1965) The functional morphology of the reproductive organs of the common (Forficula auricularia) earwig and other Dermaptera, with reference to the natural classification of the order. Journal of Zoology, 146, 1-43.
Popham, E. J. 2000. The geographical distribution of the Dermaptera (Insecta) with reference to continental drift. Journal of Natural History 34(10): 2007-2027.
Rentz, D.C.F. & Kevan, D.K.M. (1991) Dermaptera. Ed. by D. T. Naumann: Insect of Australia CSIRO, Melbourne. 360-368.
Sakai, S. (1970-1996) Dermapterorum Catalogus. Daito Bunka University.
Sakai, S. (1982) A new proposed classification of the Dermaptera with special reference to the check list of the Dermaptera of the world. Addition and Errata. Dermapterorum Catalogus, 14, 1-108.
Steinmann, H. (1986) Dermaptera: Catadermaptera I. Ed. by H. Wermuth and E. M?hn: Das Tierreich 102. Gruyter, Berlin. 345pp.
Steinmann, H. (1989) Dermaptera: Catadermaptera II. Ed. by H. Wermuth and E. M?hn: Das Tierreich 105. Gruyter, Berlin. 505pp.
Steinmann, H. (1990) Dermaptera: Eudermaptera I. Ed. by H. Wermuth and E. M?hn: Das Tierreich 106. Gruyter, Berlin. 558pp.
Steinmann, H. (1993) Dermaptera: Eudermaptera II. Ed. by H. Wermuth and M. Fischer: Das Tierreich 108. Gruyter, Berlin. 709pp.
Willmann, R. (1990) Die Bedeutung pal?ontologischer Daten f?r die zoologische Systematik. Verhandlungen der Deutschen Zoologischen Gesellschaft, 83, 277-289.
Title Illustrations

| Scientific Name | Hemimerus talpoides |
|---|---|
| Comments | Hemimerina |
| View | lateral view |
| Copyright | © 1996 Fabian Haas |

| Scientific Name | Arixenia esau |
|---|---|
| Comments | Arixenina |
| Copyright | © 1996 Fabian Haas |

| Scientific Name | Forficula auricularia |
|---|---|
| Comments | Forficulina: Forficulidae |
| Copyright | © 1996 Fabian Haas |
About This Page
I am grateful to Dr. RG Beutel, Dr. RD Briceño, Dr. G Cassis and Mr. G Simpson for critically reading the mauscript.
Fabian Haas

Institut für Spezielle Zoologie und Evolutionsbiologie, Jena, Germany
Page copyright © 1996
All Rights Reserved.
- First online 18 July 1996
- Content changed 18 July 1996
Citing this page:
Haas, Fabian. 1996. Dermaptera. Earwigs. Version 18 July 1996 (under construction). http://tolweb.org/Dermaptera/8254/1996.07.18 in The Tree of Life Web Project, http://tolweb.org/